|
|
The following article is reprinted with permission from Orvin
E. Wagner, "A Plants Response to Microgravity as a Wave Phenomenon", AIP Conference
Proceedings 504, 2000, pp. 368-373. ©2000, American Institute of Physics.
This article may be downloaded for personal use only. Any other use requires
prior permission of the author and the American Institute of Physics. Dr. Wagner
gave the invited presentation on February 2, 2000, in Albuquerque. Click
here to visit the American Institute of Physics' Conference Proceedings.
A Plant's Response to Microgravity as a Wave Phenomenon
Orvin E. Wagner
Wagner Research Laboratory, Grants Pass, OR 97527
Telephone: (541)507-1539; E-mail: oedphd@gmail.com
ABSTRACT
Simple observations of plants growing on earth indicate that plants are operated by heretofore unknown waves that interact strongly with gravity. Indications of these waves and their interactions with earth's gravity include discrete frequencies derived from internodal spacings, discrete preferred angles of growth for branches, ratios of averages of reciprocals of horizontal and vertical internodal spacings that are ratios of small integers, and linear relationships between angles of growth and means of reciprocals of cell lengths. It is proposed that the lengths of cells grown in a microgravity environment be studied to see if the mean frequencies derived from cell lengths are uniform in every direction and match frequencies derived from cells grown horizontally on earth. If this is the case, then apparently no orientation dependence exists. Thus all directions would be completely equivalent to a plant growing in a microgravity environment. The cell parameters obtained would then indicate that centripetal force and gravity are equivalent for plants grown in a microgravity environment. Also one could find out if a plant's ability to function in space is improved if the plant being tested has a minimum sensitivity to gravity on earth.
INTRODUCTION
In 1988 I discovered low velocity longitudinal waves in plants and subsequently studied their basic characteristics (Wagner, 1988-1999). These waves are called W-waves because they were first found by probing freshly cut live wood. The initial velocity found for these waves was found to be close to 1 m/s (Wagner, 1989,1990). These waves don't appear to be explicitly electromagnetic but in live materials they shift charge because charge is free to move. These waves usually appear as standing waves so that with multiple probes or probing one can often find evidence for a standing waves in plant materials with charge located in periodically spaced piles. Standing waves appear to be responsible for the placement of structures such as branches or leaves (Wagner, 1990 and subsequent papers). The standing waves also appear to have an important influence on determining the size and shape of cells and shorter wavelengths are hypothesized to be important in determining cell structure (Wagner 1999). The author is of the opinion that W-waves are really an extension of quantum waves to the macroscopic with a continuous connection to the microscopic. The waves are also found outside of plants traveling with much larger velocities (v). W-waves appear to have many unique frequencies (f). These frequencies can be measured electronically, by beating with weak electromagnetic signals (Wagner 1989, 1990) and directly with a low frequency spectrum analyzer. One can also measure plant internodal spacings (s) and use these measurements to calculate the same characteristic plant frequencies using a previously measured wave velocity (Wagner, 1990, 1996). A frequency for a particular spacing is given by f=v/2s since we assume an internodal spacing is determined by a half wavelength of a standing wave. Characteristic plant frequencies repeat from plant to plant.
For this paper the most important observation is that W- waves seem to be influenced tremendously by the gravitational field in the structure of the plant. The amount of influence seems to be dependent on the plant structure. The velocity of these waves within a plant may be different depending on whether they are traveling along the gravitational field or perpendicular to it or at angles in between (Wagner, 1996, 1997). The frequencies of the waves may also be shifted to lower values when traveling along the gravitational field as compared to traveling perpendicular to the gravitational field (Wagner 1996) (Figures 1 & 2).
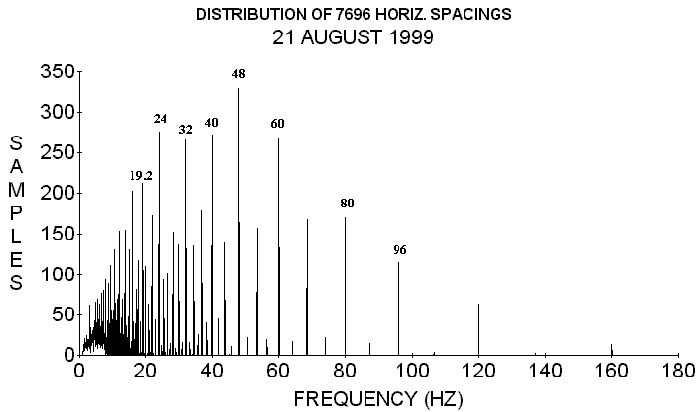
FIGURE 1. Using 96 cm/s for the velocity this is a distribution of
internodal spacing frequencies derived from 7696 horizontal internodal
spacings on plants. Compare the major peaks with those of Figure 2.
Notice that the frequencies here are considerably larger on average.
(From O. E. Wagner, Physiol. Chem. Phys. &; Med. NMR 28, 173-196 (1996).
Used by permission)
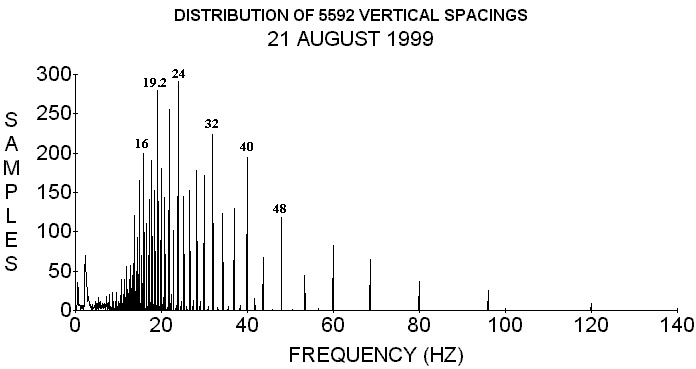
FIGURE 2. Again using 96 cm/s for the velocity this is the internodal
spacing frequency distribution of 5596 vertical internodal spacings.
Notice how the apparent frequencies are in general lower than those in
Figure 1. (From O. E. Wagner, Physiol. Chem. Phys. & Med. NMR 28,
173-196 (1996). Used by permission)
The species from which the spacings were taken, using a linear measuring instrument, in Figures 1 and 2 are Red alder (Alnus rubra), delicious apple (Pyrus Malus sp.), Himalaya blackberry (Rubus thyranthus), bracken fern (Pteridium equilinium), golden chinkapin (Castinopis chrysophylla), weeping flowering cherry (Prunus subhintella), Douglas fir (Pseudotsuga menziesii), false indigo (Amorpha fruiticosa), big leaf maple (Acer macrophyllum), Grand fir (Abies grandis), ponderosa pine (Pinus ponderosa), weeping birch (Betula pendula), Hind's willow (Salix hindsiana), golden weeping willow (Salix sp.), Dutch elm (Ulmus hollandica), and sweet corn (Zea mays sacarata). Most of these plants grew in the vicinity of the laboratory. Since spacings appear to repeat from plant to plant these graphs should include most plant frequencies except some of those with very short or very long internodal spacings.
In the wave model gravity may shift mean apparent frequencies to one third of the
horizontal (or maybe even to a smaller fraction) in live plant material. This results in
mean cell lengths and internodal spacings being up to three (or even more) times longer
parallel to the gravitational field compared to perpendicular to the gravitational field.
Thus if the gravitational field is missing or nearly so as with the microgravity the cell
is missing the reference that determines its shape, for example (Halstead and Dutcher,
1987). It also appears that plant parts grow at discrete angles to the gravitational
field as discussed in the next section.
PLANT GROWTH AT VARIOUS ANGLES TO THE GRAVITATIONAL
For several years I observed that often certain portions of branches of plants seemed to grow straight for considerable distances. In one case I found a red alder branch that grew away from the trunk for almost three meters with every portion of the branch including secondary branches almost perfectly level. In 1997 I published a paper describing the results of several hundred measurements of angles that these straight portions of branches made with the horizontal (Wagner, 1997). I found that branches seem to preferentially grow at approximately integral multiples of five degrees with the horizontal. The following graph is the result of combining all the angle data from the article plus a few more (948 total angles). The data were taken with a smart level in late June and early July (1996) when the branches were fully loaded with leaves and apparently equilibrium had been attained. Instead of drooping, forces seem to keep the branches growing straight for considerable distances. In the given cases the straight growth was for at least one half meter. 82 % of the angles of the straight portions were found to have grown within one degree of integral multiples of five degrees as the following graph demonstrates.
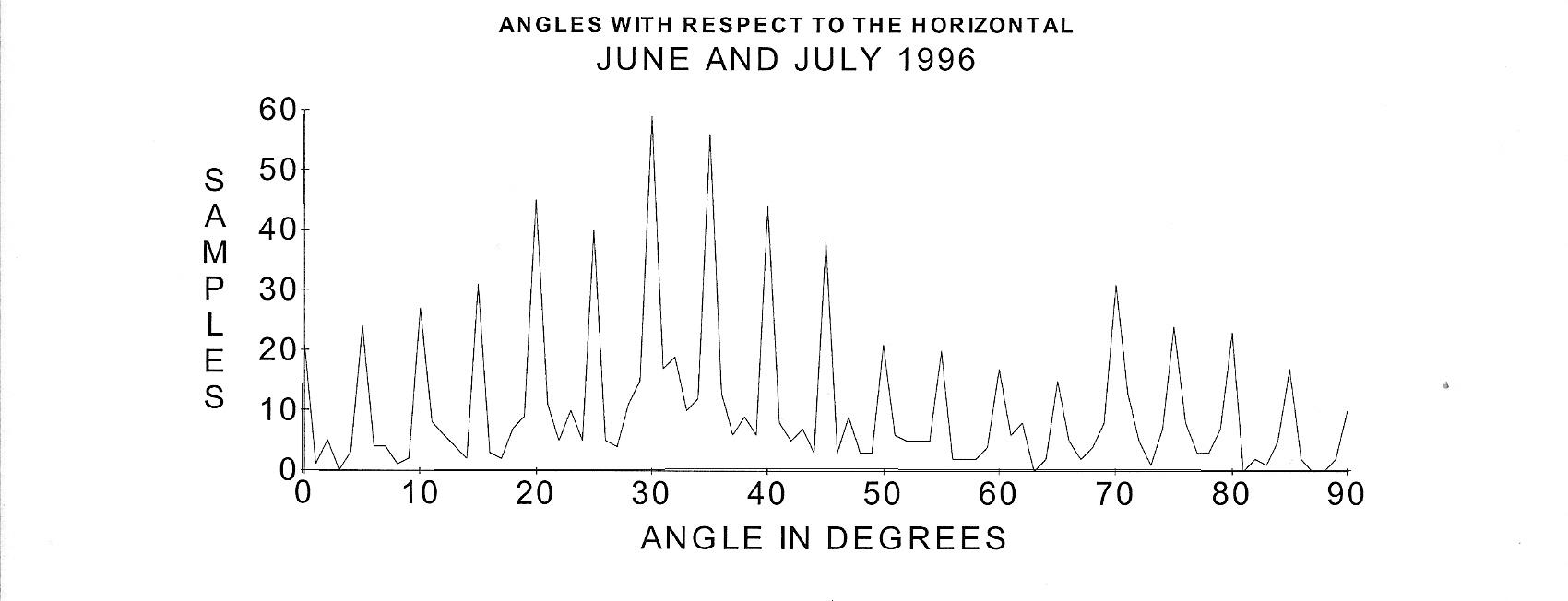
FIGURE 3. A
distribution of 948 angles of straight growth measured with respect to
the horizontal from several species of trees. The distribution
indicates that the angles of growth appear to be predominantly near
integral multiples of near 5 degrees. The data were taken from red
alder (Alnus rubra), big leaf maple (Acer macrophylla),
Douglas fir (Pseudotsuga menziesii), golden weeping willow (Salix sp.),
hazelnut (Corylus sp.), ponderosa pine (Pinus ponderosa), and an
unidentified tree species. (From O. E. Wagner,Physiol. Chem. Phys. & Med. NMR 29, 63-69 (1997). Used by permission)
The kind of behavior observed suggests
strongly that the waves involved
are interacting with gravity in a quantum like manner. The author was
aware that he could bias the measurements so he went out of his way to
be sure the data were correct by checking with a digital "smart level"
that could be read to better than one half of a degree. Care was also
taken to assure that branch roughness did not bias the measurements.
This observation of the five degree increments with respect to gravity
suggests that there are standards to correct to so that uniformity is
achieved in growing plants.
INTERNODAL SPACING AND CELL GROWTH AT DIFFERENT ANGLES TO THE GRAVITATIONAL FIELD ON EARTH
For several years I studied the ratios of
mean frequencies of internodal spacings (distances between structures
on plants such as the distances between leaves and branches). One
problem was that it is difficult to find and measure many internodal
spacings that grow at angles other than vertical and horizontal. Some
of the results from the horizontal and vertical internodal spacing data
are shown in Table 1. Here I took reciprocals of spacings multiplied by
two and then multiplied by 96 cm/s. I then found the means of the
different sets of reciprocal ratio data. 96 cm/s was used as the W-wave
velocity because it was the most commonly measured velocity in the
early work. A constant velocity was assumed for comparison purposes
even though different velocities may be present at different angles to
the gravitational field.
TABLE 1. Horizontal/vertical Internodal Frequency Ratios
Species
Number of spacings
Reciprocal ratio (H/V)
Possible velocity Ratios Big Leaf Maple
HORIZ.=205
7.10/4.81=1.48
3/2
VERT.=122
Golden Weeping Willow
HORIZ.=760
27.99/19.83=1.66
5/3
VERT.=685
False Indigo
HORIZ.=361
39.34/29.54=1.33
4/3
VERT.=376
Delicious apple
HORIZ.=618
29.99/21.68=1.33
4/3
VERT.=636
Weeping Birch
HORIZ.=380
28.35/22.71=1.25
5/4
VERT.=553
Golden chinkapin
HORIZ.=657
45.76/29.32=1.56
3/2
VERT.=373
Ponderosa Pine
HORIZ.=429
3.25/1.09=2.98
3/1
VERT.=164
Hind's willow
HORIZ.=429
57.68/39.01=1.48
3/2
VERT.=164
Red alder
HORIZ.=472
15.93/9.38=1.70
5/3
VERT.=794
Table 1 shows ratios
(H/V) of the means of reciprocals of horizontal and vertical internodal
spacings from 9 species of trees and shrubs. The ratios here could be
due to differences in velocity between vertical and horizontal or due
to difference in frequency averages as was indicated in Table 2 for
data taken from lengths of xylem cells. In Wagner 1996 it was assumed
that velocity ratios are involved because I directly measured
velocities with such ratios but the cell data given in this paper may
imply frequency ratios which was a possibility given in Wagner, 1996.
It is believed plants can utilize both different velocities and
different frequencies in gene implementation of plant structure. (From
O. E. Wagner, Physiol. Chem. Phys. Med. NMR 28, 173-196 (1996). Used by permission)
After measuring many internodal spacings, cell lengths from xylem tissue growing at specific angles to the horizontal were measured. Easy cells to measure are wood cells (tracheids, true fibers, and vessel elements) from the xylem of trees. Unlike internodal spacings many samples of cell lengths growing at different angles to the gravitational field are easy to find and analyze using a ordinary optical microscope after simple chemical separation of the xylem cells. I measured thousands of cell lengths (s) grown at different angles to the gravitational field (actually with respect to the horizontal which is the complementary angle). I again converted these reciprocals to frequency (f) using 96 cm/s (v) and then took averages of these frequencies (at least 300 for each vertical or horizontal case) . Table 2 shows the results from surface xylem cells of five different species together with the angles of growth with reference to the horizontal for the specific samplings:
TABLE 2. Mean Frequencies Versus Angle
Species
0°
5°
45°
65°
75°
80°
85°
90° Oregon ash (Fraxinus latifola)
1110
820
551
431 Pacific madrone (Arbutus menziesii)
1317
1007
877
775
Black cottonwood (Populus trichocarpa)
936
876
829
California black oak (Quercus kellogii)
642
493
420
Incense cedar (Libocedrus deccurens)
503
358
195
Table 2 gives mean
frequencies (f=v/2s) derived from cell lengths (from xylem cells)
growing at the given angles to the horizontal. One can plot almost a
straight line for each species for angle versus mean value. Again the
frequency apparently is shifted toward lower values (and/or the
velocity is changed accordingly) as the angle approaches 0 degrees with
the gravitational field. A linear relationship with angle seems to be
found here although it could be that the relationship is linear with
the sine of the angle in some cases. It would seem reasonable that the
mean of the reciprocals should change linearly with the sine of the
angle. The quantum nature of the observed angles (Wagner, 1997) may
change the relationship, however to linear with angle rather than with
sine of the angle as I would have predicted. The taking of reciprocals
seems to be necessary to obtain the linear relationships implying that
waves are involved. (From O. E. Wagner, "A Plants Response to Gravity
as a Wave Phenomenon", The J. of Grav. Physiol. 6(1) (1999). In Press. Used by permission)
It was also observed but not published that
for increasing magnitudes of angle less than zero degrees the
frequencies again decrease. For angles between zero and ninety degrees,
for the given trees, one can approximately describe the decrease in
frequency with the equation f=f0 - kq
where f is the frequency, f0 is the frequency at zero degrees, k is the
slope of the line given by the difference, the frequency at zero
degrees minus the frequency at ninety degrees, divided by 90 degrees,
for example.The given equation doesn't take into account quantization
of angles. If precise quantization is the case then one can replace the
q with 5n with n=0,1,2,3....18.
The given k could be called the gravitational sensitivity of the plant.
I will, however, define the gravitational sensitivity as simply R=H/V
where H is the mean of sample internodal spacing or sample cell length
reciprocals and V is similarly the mean of vertical internodal spacing
or cell length reciprocals. Note that ratios from spacings and ratios
from cells may be somewhat different in the same plant. The possibility
that velocities are a large function of frequency could have an
influence here and in the finding of apparent multiple velocities
(Wagner, 1996).
CELL GROWTH AT DIFFERENT ANGLES TO THE VERTICAL IN THE MICROGRAVITY FIELD
In the microgravity in a spacecraft the
gravitational field is essentially canceled because of the centripetal
field. This suggests that the only permitted frequencies for growth in
microgravity
are the ones that pertain to horizontal growth on earth. Also the
angular quantization would be expected to be missing. The result is
that the plant is constrained to grow uniformly in every direction as
far as the gravity and centripetal fields are concerned if the
principle of equivalence applies here. It is interesting to observe
that the centripetal field and the gravity field, so far, have been
considered equivalent to plants for W-waves. This is true with the
principle of equivalence but to me this puts the idea of gravity being
a curvature of space phenomenon as Einstein assumed in doubt because
curvature of space does not seem to explain inertia or momentum. The
study of a plants response to gravity may
open up new ways of looking at gravity. For example perhaps gravity is
a very short wavelength wave phenomenon.
Almost everyone has assumed that plants
react to centripetal force and gravity in the same manner (Salisbury
1999). This may not be exactly true so perhaps measurements from plants
grown in a microgravity environment would clear up the matter. As an
initial experiment we would like to determine if any cell shape
anisotropy as a function of orientation can be detected at all in a
microgravity environment. One would have to use the proper lighting to
prevent growth anisotropy due to lighting. If uniform lighting can be
achieved then one can check if any possible difference in effect can be
found between centripetal forces and ordinary gravitational forces on
earth. One needs to be sure that all the plant interactions that
pertain to gravity are known. This may require more measurements. The
process requires taking measurements similar to the above for wood and
other cells grown in the microgravity field. There likely would not be
a large enough sampling of internodal spacings to measure on plants
grown in a microgravity environment. Plant cells, however, with their
many different orientations should be available in
abundance for measurement as long as the particular plant chosen will
grow in the microgravity environment. One should probably choose woody
plants that are proven to be sensitive to gravity from a large earthly
horizontal reciprocal to vertical reciprocal ratio (R). It is my
hypothesis that plants with a small R would be more likely to grow well
in a microgravity environment, however. This is a hypothesis that could
be tested simultaneously in the experiment. In some plants (such as
black cottonwood in table 2) the ratio between horizontal reciprocal
and vertical reciprocal averages was found to be close to one. All
plants so far measured on earth always show at least a small ratio
which is greater than one. The results would be related to the
principle of equivalence, gravitational theory, and plant-gravity
interactions.
CONCLUSIONS AND OBSERVATIONS
The wave approach to a plant's
interaction with gravity appears to answer many of the questions about
how plants are influenced by the gravity and microgravity fields. With
the additional knowledge about how a plant grows and responds to
gravity we may be able to alter the environment to produce different
types of desired growth. Perhaps one can generate the proper fields to
produce an environment that will allow plants to grow as if they were
on earth while in a microgravity environment. If the wave theory
applies to plants it likely applies in subtle ways to other life.
Likely cell growth of all kinds is influenced, to at least some extent,
everywhere, by the direction and magnitude of the gravitational field.
The subtle influences are likely important to man's survival at any
location in space.
REFERENCES
Halstead, T.W. and Dutcher, F.R., "Plants in Space", Ann. Revs. of Plant Physiol. 38, 317-345 (1987).
Salisbury, F. B., "Gravitropism: Changing Ideas", Horticultural Revs. 15, 233-278 (1993).
Salisbury, F. B., Private communication., (1999).
Wagner, O.E., "Acceleration Changes within Living Trees", Physiol. Chem. and Phys. & Med. NMR 27, 31-44 (1994).
Wagner, O.E. , "Acceleration Changes within Plants", Physiol. Chem. and Phys. & Med. NMR 24, 29-33 (1992).
Wagner, O. E., "Anisotropy of Wave Velocities in Plants: Gravitropism", Physiol. Chem. Phys. & Med. NMR 28, 173-196 (1996).
Wagner, O.E.,"Quantization of Plant Growth Angles With Respect to Gravity", Physiol. Chem. Phys. & Med. NMR 29, 63-69 (1997).
Wagner, O.E., "Wave Behavior in Plant Tissue", Northw. Sci. 62, 263-270 (1988).
Wagner, O.E., "Wave Energy Density in Plants", Physiol. Chem. and Phys. & Med. NMR 25, 49-54 (1993).
Wagner, O.E., "Waves in Dark Matter", Physics Essays. In Press (1999).
Wagner, O.E., Waves in Dark Matter, Rogue River, Wagner Physics Publishing, 1995, pp. 1-188.
Wagner, O.E., "W-waves and Plant Communication", Northw. Sci. 63, 119-128 (1989).
Wagner, O.E., "W-waves and Plant Spacings", Northw. Sci. 64, 28-38 (1990).
| Copyright ©2000-2018 Wagner Research Laboratory |
 Waves in Dark Matter
Waves in Dark Matter